SLIDE 3 3
Attainment of the equilibrium allele frequency given selection and a variety of different mutation rates. Note that the time to equilibrium varies in addition to the actual equilibrium frequencies.
s = 0.1 (Waa = 0.9) µ = mut rate A → a µ = 0.01 µ = 0.001 µ = 0.0001 µ = 0.00001
a Note that fore realistic mutation rates, the equilibrium frequencies are quite low (freq of a allele > 0.05). In this example selection pressure is also quite weak (s = 0.1). If we assume stronger selection pressure (s > 0.1), the equilibrium point will be lower and the rate to equilibrium will be faster.
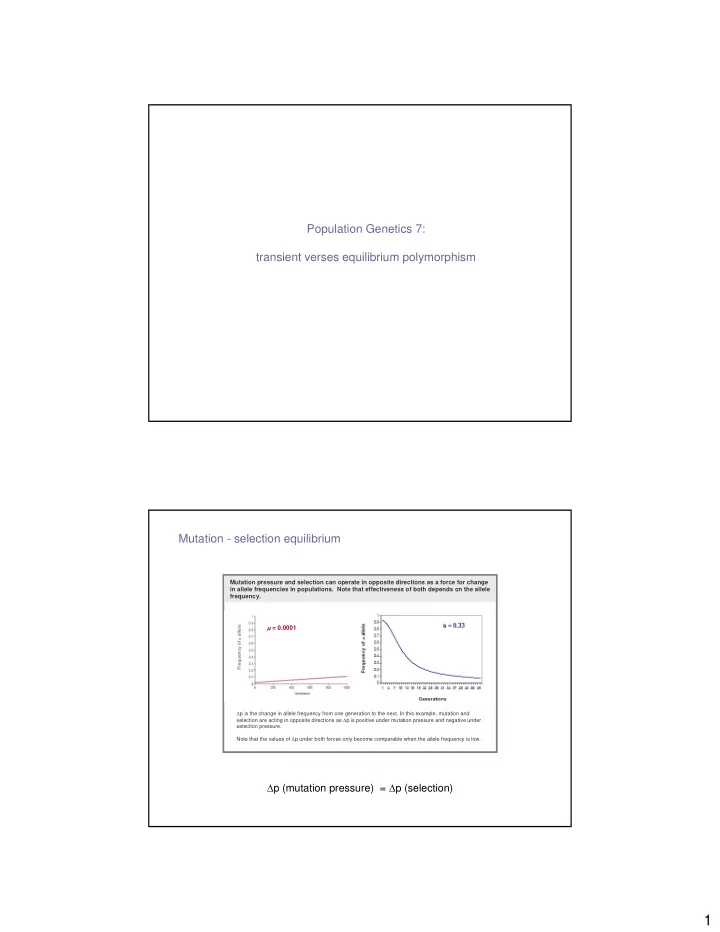
Mutation - selection equilibrium
Effect of partial dominance on mutation-selection equilibrium. The fitness of genotypes AA, Aa, and aa are assumed to be 1, 1–hs, and 1- s respectively.
0.005 0.01 0.015 0.02 0.025 0.03 0.035 0.000001 0.00001 0.0001 0.001 h = 0 h = 0.01 h = 0.05 h = 0.1 h = 0.5
Equilibrium frequency of recessive allele (a) Ratio of mutation rate to selection coefficient against aa (µ/s) The symbol h is the amount of dominance in the heterozygote genotype. Note, that even a small amount of dominance (h = 0.01) reduced the equilibrium frequency of the recessive
- allele. Hence, dominance has a significant influence on the equilibrium point. The reason
is that when q, the freq of the recessive allele is small, the majority of those alleles are in the heterozygote configuration, and even a small amount of selection on the heterozygotes leads to a major reduction in its equilibrium frequency as compared with full dominance. Note that for reasonable values of µ, h, and s, the equilibrium frequencies are < 0.01, This means that mutation selection equilibrium is not sufficient to explain low frequency detrimental alleles in populations where those alleles have frequencies > 0.01
Mutation - selection equilibrium