SLIDE 4 11/2/2011 4
Learned changes in the VOR in monkeys outfitted with magnifying lenses.
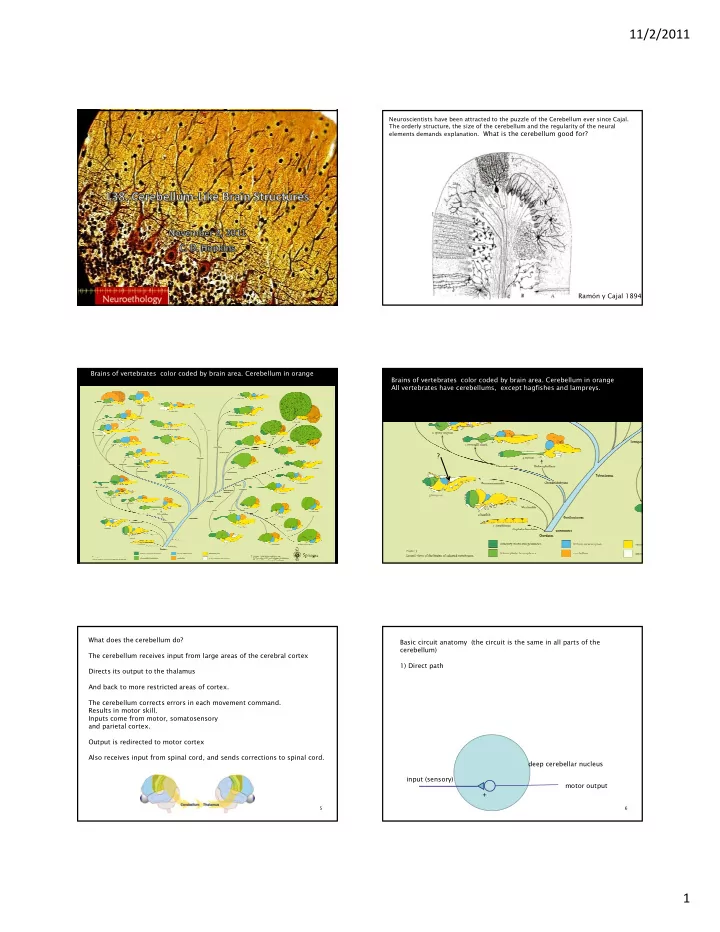
Ito’s hypothesis
Visual input from superior colliculus to inferior olive: information about retinal ‘slip’ Climbing fiber input to Purkinje cell – error signal. Vestibular input from mossy fiber to granule cell to the parallel fiber in flocculo- nodular lobe of cerebellum. When co-active with error, synaptic weight of parallel fiber synapse is weakened by long term depression. Inhibition decreases and this strengthens the drive from the vestibular system increasing the gain.
Cerebellum Function: DIRECT PATHWAY: the vestibular system to the eye muscles (VOR) INDIRECT PATHWAY: vestibular collateral (mossy fiber) granule cell parallel fiber Purkinje cell (inhibits VN). ERROR SIGNAL: error in the VOR, information about image slip on the retina comes from the climbing fiber which originates in the inferior olive. (strong depolarization) Climbing fiber causes dendritic spike in the Purkinje cell s causes synaptic weights from parallel fibers to depress, weakening the inhibitory input on the vestibular neurons, thereby adjusting the gain of the eye reflex.
The Basic Architecture
1. Bell, C. C., Han, V. and Sawtell, N. B. (2008). Cerebellum-like structures and their implications for cerebellar function. Annu Rev Neurosci 31, 1-24.
What is a Cerebellum-like structure?
Most vertebrates possess both a cerebellum and other brain structures with cerebellum-like architectures. 1) A MOLECULAR LAYER with many parallel fibers + dendrites of Purkinje-like cells. 2) A LARGE NUMBER OF GRANULE CELLS: which carry information from many central structures about the state of the
molec deep
central structures about the state of the animal: sensory information from a large number of senses, corollary discharges from motor centers (predictive of sensory input to the principle cells). 3) PRINCIPAL CELLS with SPINES. Parallel fibers terminate on spines of principal cells; or on the smooth dendrites of inhibitory stellate cells.
Cerebellum-like structures sometimes serve as adaptive sensory filters that predict sensory inputs into the deep layers using the associated parallel fiber inputs in the molecular layer.
The cerebellar-like structure is defined by the molecular layer, the principal cell, and the numerous inputs from granule cells via parallel