
SLIDE 1
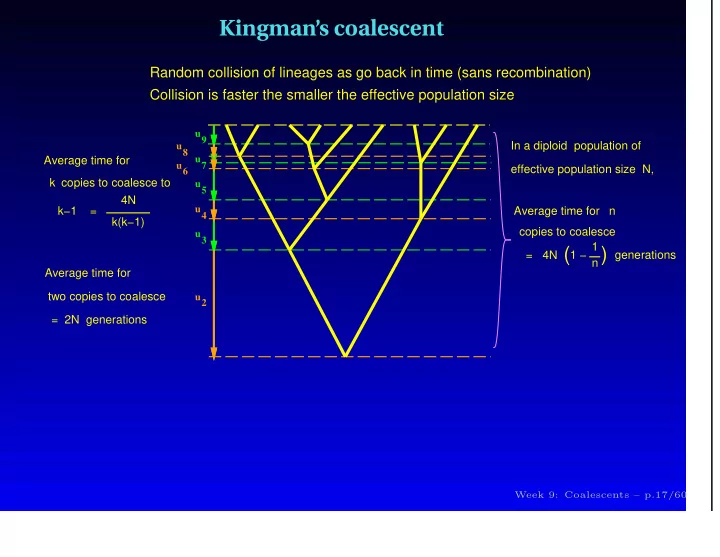
Kingman’s coalescent
u9 u7 u5 u3 u8 u6 u4 u2
Random collision of lineages as go back in time (sans recombination) Collision is faster the smaller the effective population size
Average time for n Average time for copies to coalesce to 4N k(k−1) k−1 = In a diploid population of effective population size N, copies to coalesce = 4N (1 − 1 n
(
generations k Average time for two copies to coalesce = 2N generations
Week 9: Coalescents – p.17/60