
SLIDE 1
Linking phytoplankton phenology to pink salmon productivity along a north-south gradient Michael J. Malick1, Sean P. Cox1, Franz J. Mueter2, and Randall M. Peterman1
1 School of Resource and Environmental Management, Simon Fraser University, Burnaby, British
Columbia, Canada
2 School of Fisheries and Ocean Sciences, University of Alaska Fairbanks, Juneau, Alaska, USA
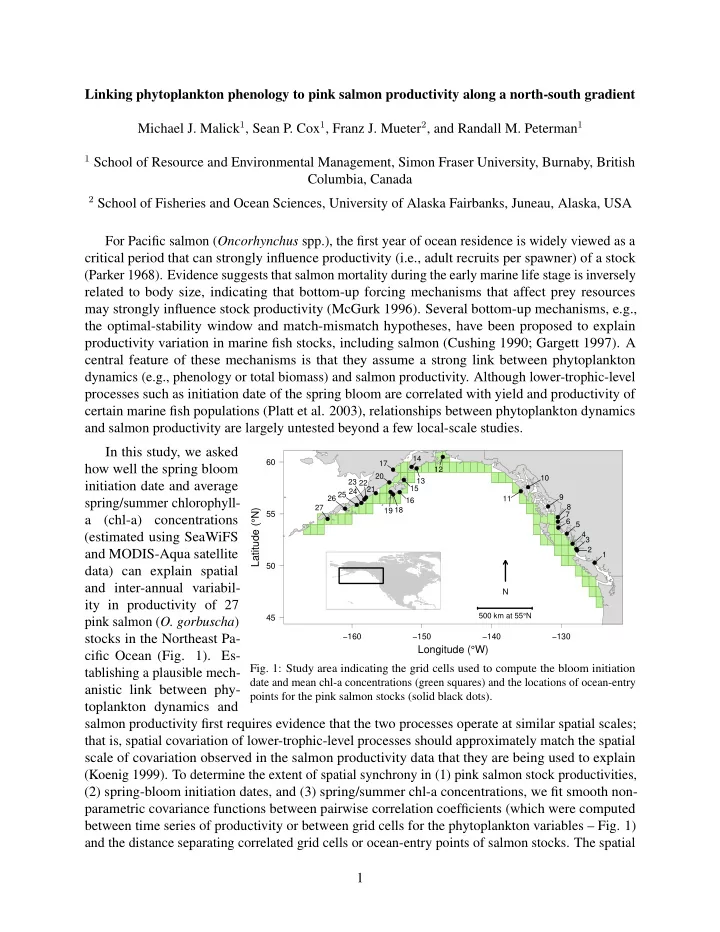
For Pacific salmon (Oncorhynchus spp.), the first year of ocean residence is widely viewed as a critical period that can strongly influence productivity (i.e., adult recruits per spawner) of a stock (Parker 1968). Evidence suggests that salmon mortality during the early marine life stage is inversely related to body size, indicating that bottom-up forcing mechanisms that affect prey resources may strongly influence stock productivity (McGurk 1996). Several bottom-up mechanisms, e.g., the optimal-stability window and match-mismatch hypotheses, have been proposed to explain productivity variation in marine fish stocks, including salmon (Cushing 1990; Gargett 1997). A central feature of these mechanisms is that they assume a strong link between phytoplankton dynamics (e.g., phenology or total biomass) and salmon productivity. Although lower-trophic-level processes such as initiation date of the spring bloom are correlated with yield and productivity of certain marine fish populations (Platt et al. 2003), relationships between phytoplankton dynamics and salmon productivity are largely untested beyond a few local-scale studies.
−160 −150 −140 −130 45 50 55 60
Longitude (°W) Latitude (°N)
- ●
- ●
- ●
- 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 N 500 km at 55°N
- Fig. 1: Study area indicating the grid cells used to compute the bloom initiation