
1
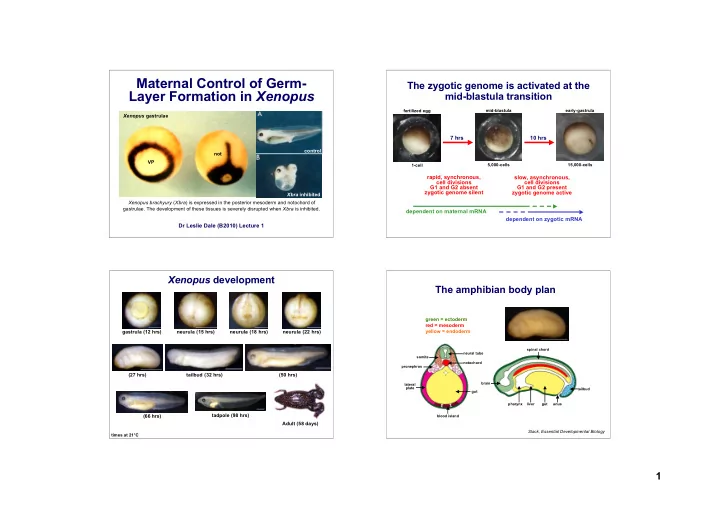
Maternal Control of Germ- Layer Formation in Xenopus
Dr Leslie Dale (B2010) Lecture 1
Xenopus brachyury (Xbra) is expressed in the posterior mesoderm and notochord of
- gastrulae. The development of these tissues is severely disrupted when Xbra is inhibited.
VP not Xenopus gastrulae control Xbra inhibited
The zygotic genome is activated at the mid-blastula transition
rapid, synchronous, cell divisions G1 and G2 absent zygotic genome silent slow, asynchronous, cell divisions G1 and G2 present zygotic genome active dependent on maternal mRNA dependent on zygotic mRNA 7 hrs 10 hrs
1-cell fertilized egg 5,000-cells mid-blastula 15,000-cells early-gastrula
Xenopus development
times at 21°C
gastrula (12 hrs) neurula (15 hrs) neurula (18 hrs) neurula (22 hrs) (27 hrs) tailbud (32 hrs) (50 hrs) Adult (58 days) tadpole (98 hrs) (66 hrs)
The amphibian body plan
Slack, Essential Developmental Biology
neural tube notochord somite pronephros lateral plate blood island brain spinal chord pharynx liver gut anus tailbud gut
green = ectoderm red = mesoderm yellow = endoderm